|
Fondazione FONAMA |
fonama@fonama.org |
Tel. 335250742 |
 |
 |
Traduzioni a cura di Natale Marzari
Dopo 41 anni e 5 mesi, nel maggio 2006 la
magistratura di Trento ha riconosciuto l'esistenza e la gravità di quella
malattia rara che nessuna altra istituzione o persona singola della provincia di
Trento ancora mi riconosce, e per negare la quale ancora mi perseguita.
Natale Marzari |
Agenti disaccoppianti e inibitori
Molta della nostra conoscenza della funzione mitocondriale
deriva dallo studio delle sostanze tossiche. Gli inibitori specifici sono stati
usati per distinguere il sistema di trasporto
dell'elettrone dal sistema di
fosforilazione ed hanno aiutato a definire la sequenza degli
elementi portatori di riduzione lungo
la catena respiratoria. Se la catena è bloccata allora tutti gli intermediari
dal lato del
substrato del blocco sono ridotti, mentre tutto quelli dal lato dell'ossigeno
sono ossidati. È facile da vedere che cosa è accaduto perché gli elementi
portanti
ossidati e ridotti differiscono spesso nelle loro proprietà
spettrali. Se è disponibile una gamma di inibitori diversi allora molti portatori respiratori possono essere disposti nell'ordine
corretto.
Ci sono sei tipi distinti di sostanze tossiche che possono interessare la
funzione mitocondriale:
1) Inibitori della catena respiratoria (per es. cianuro, antimicina, rotenone
& TTFA) bloccano la respirazione in presenza sia di ADP sia di disaccoppiatori.
2) Inibitori della fosforilazione (per es, l'oligomicina) abolisce
la punta di
consumo dell'ossigeno dopo l'aggiunta dell'ADP, ma non ha effetto sulla respirazione stimolata
da disaccoppiatore.
3) Agenti di disaccoppiamento (per es. dinitrofenolo, CCCP, FCCP) aboliscono il
collegamento obbligatorio fra la catena
respiratoria ed il sistema di fosforilazione che si osserva nei
mitocondri intatti.
4) Inibitori di trasporto (per es. atrattiloside, acido bongcrecico, NEM)
sia prevenendo l'esportazione dell?ATP, sia l'importazione delle
materie prime attraverso la membrana interna
mitocondriale.
5) Ionofori (per es. la valinomicina, il nigericin) rendono membrana interna
permeabile a sostanze che ordinariamente non
possono attraversarla.
6) Inibitori del ciclo di Krebs (per es. l'arsenito,
l'aminoossiacetato) i quali bloccano uno o più degli enzimi del ciclo
TCA, o una reazione collegata.
Alcune delle sostanze più note sono elencate qui sotto:
| Sostanze |
Modo di azione ed effetti
|
|
Ossiacetato Amminico |
Inibitore degli enzimi secondari. Inibisce tutte le transaminasi reagendo in
covalenza con il loro
gruppo prostetico fosfato piradossale. Blocca il ciclo malato/aspartato
inibendo glutammato - ossaloacetato transaminasi (GOT) |
| Antimicina A |
Inibitore della respirazione. Blocca la catena respiratoria al complesso
III fra
il citocromo b ed
il citocromo c1. Quindi impedisce l'ossidazione sia del NADH che del succinato,
ma non ha effetto sull'ascorbato + TMPD. |
| Arsenito |
Inibitore del ciclo di Krebs. Reagisce con il legame fra
disolfuro e l'acido lipoico ossidato, formando un complesso ciclico. Inibisce tutte le
deidrogenasi osso-acide, compresa la piruvato deidrogenasi,
la ossoglutarato deidrogenasi e la deidrogenasi della catena ramificata osso-acida. |
| Atrattilosido
|
Inibitore di trasporto. Bloccail portatore del nucleotide adenina
legandosi alla faccia
all'esterna della formazione (contrasto con acido
bongcrecico). Non ha effetto sulle particelle sub-mitocondriali, le quale si risigillano spontaneamente
con le membrane esterne dopo l'ultrasonificazione. Questo inibitore del
trasporto ATP/ADP assomiglia all'oligomicina quando si usa con i mitocondri intatti. (vedi inoltre l'acido di Bongcrecico.) |
| Acido Bongcrecico |
Inibitore di trasporto. Blocca il portatore del nucleotide adenina
legando ai
rivestimenti della conformazione interna (contrasto con il
atrattilosido). |
| Cianuro |
Inibitore della respirazione. Blocca la citocromo ossidasi (complesso
IV) ed
evita sia la
respirazione accoppiata che disgiunta con tutti i substrati, compreso la NADH,
il succinato e l'ascorbato + TMPD. |
| Mersalile |
Inibitore di trasporto. Dose d'inibizione dipendente dai portatori del dicarbossilato e del
fosfato. Il mersalil è un diuretico mecuriale antiquato. |
| maleimide dell'N-etile (NEM) |
Inibitore di trasporto. Blocca il portatore fosfato reagendo con i
gruppi SH, ed impedisce la respirazione accoppiando mitocondri e fosfato -
gonfiamento mediato. |
| Oligomicina |
Inibitore della fosforilazione. Si lega ad un polipeptide23kd (OSCP) della piastra di base F0 e
blocca la sintesi dell'ATPi (e
degradazione) dalla F0/ATPasi F1. Abolisce la respirazione stimolata dall'ADP
nei
mitocondri intatti ed in
tutte le funzioni alimentate dall'ATP nelle particelle sub-mitocondriali. |
| Rotenone
|
Inibitore della respirazione. Blocca la deidrogenasi NADH (complesso
I) nella
catena respiratoria
ma non ha effetto sull'ossidazione sia del succinato sia dell'ascorbato + di
TMPD. (TMPD è tetra diammina metilica di fenilene, un mediatore redox
artificiale che aiuta il trasferimento degli
elettroni dall'ascorbato al citocromo c.) |
| Acetone trifluorotenolo (TTFA) |
Ostruisce la succinato deidrogenasi (complesso II). Non ha effetto
sull'ossidazione dei substrati collegati alla NADH sia dell'ascorbato +
TMPD |
Disaccoppiatori e
ionofori: Tutte queste sostanze sono piccole molecole amfipatiche
che si sciolgono nel doppio foglio fosfolipidico ed aumentano enormemente la
sua permeabilità ionica. Proteggono la carica elettrica mentre lo ione
attraversa la membrana, fornendo un ambiente polare per lo ione e una faccia
idrofoba al mondo
esterno. L'ionoforo trasporta una varietà di ioni, ma disgiungendo specificamente
gli agenti aumenta la permeabilità protonica, e sconnette la catena di trasporto degli elettrone dalla formazione
dell'ATP. Alcuni ionofori sono prodotti naturali isolati dai
microorganismi, ma altri sono sostanze sintetiche, adeguate ad un'applicazione
specifica. Gli agenti disgiungenti sono tutti sintetici, anche se ci è regolato
delicatamente disgiungendo la proteina addetta alla termogenesi
nei mitocondri marrone del tessuto adiposo.
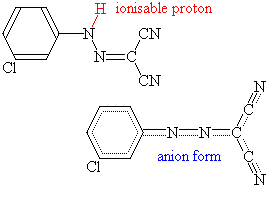 |
CCCP: (idrazone fenilico m.-cloro del cianuro di carbonile) questo è un
acido debole lipido-solubile ed è un agente disgiungente
mitocondriale molto potente. Il FCCP (idrazone fenilico del cianuro di carbonile di
p-trifluoromethossile) è simile.
La carica negativa è ampiamente delocalizzata su circa dieci
atomi nella forma ionizzata di CCCP, così il campo elettrico che circonda
l'anione di CCCP è molto
debole. Ciò permette che l'anione si diffonda liberamente con i mezzi non
polari, quali le membrane del fosfolipide. Questo comportamento è molto
insolito: la vasta maggioranza degli ioni elettricamente caricati è esclusa
dagli ambienti non polari.
|
Nei mitocondri intatti, il CCCP entra in formaprotonata, scarica
il gradiente pH, ed allora agisce subitoi come anione, distruggendo il potenziale
di membrana. Il processo può essere ripetuto milioni di volte, in modo che una
quantità molto piccola di CCCP può catalizzare il
movimento di un numero enorme di protoni, e cortocircuitare la catena respiratoria.
L'ionoforo può essere diviso in forma di canale
(tipo la gramicidina) che formano un poro molto piccolo attraverso la
membrana, e gli elementi portanti mobili che si diffondono avanti ed indietro
attraverso la membrana. Gli ionofori sono usati studiare i mitocondri
normalmente
appartengono al gruppo dei portatori mobili. Mostrano una specificità
ionica considerevole perché lo ione deve
essere accomodato all'interno di uno spazio limitato all'interno del
portatore:: ci sono ionofori del potassio, ionofori del calcio e
così via.
Valinomicina: Questo portatore mobile catalizza il movimento elettrico del K+
attraverso il doppio foglio fosfolipidico. Ciò implica che la distribuzione del
potassio attraverso la membrana obbedisca all'equazione
di Nernst una
volta che è stato raggiunto l'equilibrio.
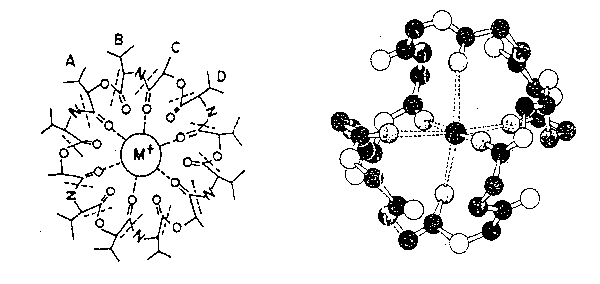
È un'ammide ciclicaestere nel quale la sequenza isovalerato D-idrossilato, L-valina, L-allatti e la D-valina è ripetuta tre volte (- A-B-C-D-
nello
schema). Lo ionoforo fornisce un polare interno per ospitare lo ione
potassio, ma presenta un esterno lipofilico non polare al mondo esterno.
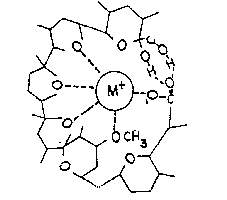 |
Nigericina: Questo portantore mobile assomiglia alla valinomicina ma contiene
un gruppo carbossilico e forma un sale del potassio. Quindi catalizza e
neutralizza elettricamente un potassio/protone scambiato attraverso il doppio
foglio fosfolipidico. Ciò implica che la distribuzione del potassio sia collegata con
l'equilibrio del gradiente pH una volta sia stato raggiunto:
[K+ ]in / [H+ ]in = [K+
]out / [H+ ]out
|
In alcuni casi (per es. la valinomicina) lo ionoforo - ione complesso ha
una carica
elettrica netta, considerando che l'elemento portante vuoto è neutro. Altri
ionofori (per es. la nigericina) forma soltanto complessi elettricamente neutri.
La carica fa una differenza enorme al comportamento di ogni elemento
portante, poiché i complessi caricati interagiscono con tutti i potenziali
della membrana, considerando che i complessi neutri rimangono inalterati. Parliamo
dei meccanismi elettrici ed elettricamente neutri
dell'elemento portante per attirare l'attenzione su questa differenza.
Questo concetto può estendersi per coprire le proteine naturali di
trasporto nella membrana mitocondriale interna. Molte di queste proteine (ed
alcuni ionofori)
catalizzano il passaggio di un legando per un altro, quindi il termine antiportatori.
Inoltre parliamo degli simportatori (due molecole
che viaggiano insieme nello
stesso senso) e di uniportatori (una molecola viaggia da sola.) ancora una volta, la
precisa della carica stechiometrica ha un'influenza enorme sui
risultati.
Termogenesi da
brivido: I mitocondri che si trovano nel tessuto adiposo marrone
contengono una
proteina disgiungente unica denominata termogenina, la quale permette l'entrata
controllata dei protoni senza stesi di ATP per generare calore. La proteina è un dimero 33kd,
relativo strutturalmente al portatore nucleotide
adenina. È inibito da
GTP, e l'attivazione è controllata dal sistema nervoso simpatico. Questo
processo è particolarmente importante in bambini appena nati, quale può perdere
molto velocemente il calore ai loro dintorni, ma si presenta inoltre negli adulti
ed in animali in ibernazione. Contrariamente al tessuto adiposo bianco più
familiare,
il grasso marrone ha un rifornimento capillare eccellente di sangue e
può realizzare i tassi metabolici molto alti. Il deposito principale in esseri
umani è dietro le scapole, con altre zone lungo la spina dorsale. Il colore marrone risulta dagli enzimi
respiratori.